(I'm flirting with starting a running "people don't understand..." theme on the blog, considering the endless supply of topics it supplies.)
There is a general public awareness, at least in the United States, that there exists this thing called "The Liberal Media". To a certain extent, that label can be justified. Journalism, as a profession, does tend to be populated by people who identify as liberals much more than conservatives. Unfortunately, that isn't how the term is usually used. People aren't claiming there are liberals in the media... they're claiming the content of the media is deliberately and blatantly biased towards liberal ideology.
Now here's the problem. The people making this claim have a definition of "biased" that is... well... let's call it "divorced from reality". You see, bias in journalism is when instead of reporting the plain facts of the matter at hand, you distort them to say something else that suits your own agenda. In the minds of the people who rant and rave about "The Liberal Media" however bias is when you report a bad thing about someone they like, and don't immediately follow it with either a good thing about someone they like, or a bad thing about someone they don't like. Because see... you have to be "balanced".
Now I may be crazy, but I always thought the job of the news was to report the facts, not to play reality referee and dishonestly manipulate the reporting of the facts so it looks like both sides of any ideological dispute were perfectly equal. I'm pretty sure in fact that THAT would be displaying bias.
For an example of what we're talking about, this startlingly insightful individual (you'll encounter their like a lot if you spend any amount of time in political discussion forums) has concluded that Fox News provided the fairest news coverage in the country of the 2008 elections.
Yeah, you're reading the title of the link right. Fox. When you click on that link you'll find the study they cite to back up this entertaining claim is one done by the Pew Research Center's Project for Excellence in Journalism which... counted how many positive and negative things different networks said about the candidates. Seriously.
"Well... they said this many nice things about one side... but only this many nice things about the other side... BIAS!"
Yes, the only possible way that could happen is biased reporting. As opposed to... just possibly... there actually being in real life more positive or negative aspects to report about one side than the other. (But nah... it's clear McCain ran exactly as effective a political campaign as Obama did, right? I mean Obama ran what is widely regarded as one of the most effective campaign operations in modern political history... and McCain told the nation that Sarah Palin was the most qualified person in America to run the country after him. That's got to balance out, right?)
You know, there are days when it's really hard to be optimistic about the future of the species.
Let's apply these brilliant criteria for unbiased journalism to a hypothetical test case. We're going to travel back in time and cover, say, one of the trials of Charles Manson. On the one hand, we have the coverage by GNN (Generic New Network). On the other hand, we have the coverage by Faux News.
GNN: Every story they write is talking about this guy being accused of murdering people! They mention he's a repeat offending ex-convict! It's all negativity all the time!They never say anything nice about the guy!!!
Faux: They put up some stories about him being accused of murdering people. But they also put up just as many stories about how he considers himself a dedicated family man.
GNN is, therefore, 100% horribly horribly biased. Faux News on the other hand is what journalism is meant to be. They're balanced. And that's fair.
And that's what's really important in journalism. Right?
5.31.2010
5.30.2010
Proving Evolution : Post 7 - Piling On... And Finishing Up.
Chromosome Fusions
A lot of animals have different numbers of chromosomes. An often raised objection to evolution is that this means at some point an organism would have been born with a different number of chromosomes from the rest of the population but it wouldn’t have had anything it could mate with that had the same number of chromosomes so the mutation wouldn’t have been preserved. This objection is based on the false idea that animals with different numbers of chromosomes are incapable of interbreeding.
If this was true the existence of modern domesticated horses would be something of a genetic miracle. Domestic horse populations have 64 chromosomes… wild horse populations have 66.
In reality chromosome fissions and fusions are hardly an unknown phenomenon.
One such fusion clearly occurred after the hominids branched off from the rest of the primates. Humans have 23 pairs of chromosomes, all the rest of the primates have 24. Evolutionary theory and the nested hierarchy then tells us this means there was a fusion event which reduced the number of chromosomes in humans to 23 after their ancestors split off from the wider population. If this prediction is true, we should be able to see clear evidence of it in a chromosomal analysis.
Lo and behold:
Chromosome fusion
There is overwhelming evidence that human chromosome 2 is the product of the fusion of two chromosomes which just happen to look basically identical to two chromosomes found in chimpamzees… as seen in the image included in the above link.
Note that this is not just evidence that human and chimp genetic sequences kind of look the same. The telomere and centromere sequences in the middle of human chromosome 2 are clear indication that that chromosome is the product of the combination of two different pre-existing independent chromosomes. If humans had been independently created in their modern form rather than having evolved into it from a common ancestor with other animals there is no reason to expect find something like this in the human genome… but there it is.
Biogeography and Paleobiogeography
Biogeography is the mapping of spatial patterns of biodiversity. Ie: which animals and types of animals are found in which geographic regions. Combined with paleobiogeography, which is the mapping of the same in the fossil record, this presents us with yet another piece of corroborating evidence for evolution. Fossil forms which are morphologically transitional stretching back from modern animals back to earlier ones are found in geographically contiguous locations throughout the record. Obviously this is something which is to be expected if all those transitional forms were to have evolved one from the other. If they were not transitional ancestral organisms but rather just completely independent separately created lineages of some kind there would be no reason to expect the geographical distributions we do observe that they fall into.
Properties of DNA Replication
A lot of animals have different numbers of chromosomes. An often raised objection to evolution is that this means at some point an organism would have been born with a different number of chromosomes from the rest of the population but it wouldn’t have had anything it could mate with that had the same number of chromosomes so the mutation wouldn’t have been preserved. This objection is based on the false idea that animals with different numbers of chromosomes are incapable of interbreeding.
If this was true the existence of modern domesticated horses would be something of a genetic miracle. Domestic horse populations have 64 chromosomes… wild horse populations have 66.
In reality chromosome fissions and fusions are hardly an unknown phenomenon.
One such fusion clearly occurred after the hominids branched off from the rest of the primates. Humans have 23 pairs of chromosomes, all the rest of the primates have 24. Evolutionary theory and the nested hierarchy then tells us this means there was a fusion event which reduced the number of chromosomes in humans to 23 after their ancestors split off from the wider population. If this prediction is true, we should be able to see clear evidence of it in a chromosomal analysis.
Lo and behold:
Chromosome fusion
There is overwhelming evidence that human chromosome 2 is the product of the fusion of two chromosomes which just happen to look basically identical to two chromosomes found in chimpamzees… as seen in the image included in the above link.
Note that this is not just evidence that human and chimp genetic sequences kind of look the same. The telomere and centromere sequences in the middle of human chromosome 2 are clear indication that that chromosome is the product of the combination of two different pre-existing independent chromosomes. If humans had been independently created in their modern form rather than having evolved into it from a common ancestor with other animals there is no reason to expect find something like this in the human genome… but there it is.
Biogeography and Paleobiogeography
Biogeography is the mapping of spatial patterns of biodiversity. Ie: which animals and types of animals are found in which geographic regions. Combined with paleobiogeography, which is the mapping of the same in the fossil record, this presents us with yet another piece of corroborating evidence for evolution. Fossil forms which are morphologically transitional stretching back from modern animals back to earlier ones are found in geographically contiguous locations throughout the record. Obviously this is something which is to be expected if all those transitional forms were to have evolved one from the other. If they were not transitional ancestral organisms but rather just completely independent separately created lineages of some kind there would be no reason to expect the geographical distributions we do observe that they fall into.
Properties of DNA Replication
Proving Evolution: Post 6 - Phylogenetic Analysis
Previously we skimmed over the creation of a phylogenetic tree with a simplified example of how they are constructed using only a few major genetic characteristics. In the last post we touched on how even much less obvious genetic characteristics can also be analyzed for phylogenetic relationships… like ERVs. As the discussion progresses the importance of the nested hierarchy and it’s nontrivial nature will continue to become more apparent. Like in the case of ERVs it goes significantly beyond such superficially obvious observations as “we never expect to find snakes producing orange juice”. It applies right down to the molecular level even to genetic sequences which have absolutely no reason, from the standpoint of observing the “obvious” groupings of organisms, to display nested hierarchical patterns... except that evolutionary theory says they should because of their patterns of common ancestry.
When actually constructing a consensus phylogenetic tree such as the one shown at (Life on Earth) not only are a great many genetic traits taken into account, but a rigorous mathematical analysis of the actual DNA sequences of the organisms in question (where such DNA is available) is done to create cladograms (the branching diagrams showing patterns of descent) with the highest possible percentage confidence. These techniques have been tested in situations where the correct evolutionary relationships are already independently known for an absolute certainty to verify that they do in fact not simply produce an evolutionary relationship but the correct evolutionary relationship to within a very low margin of error..
One example:
http://mbe.oxfordjournals.org/cgi/reprint/19/2/170.pdf
In the paper above the researchers started with an original sample of DNA from Trypanosoma cruzi. They bred it over successive generations and allowed it to continually mutate, and every 70 generations 2 of the resulting DNA sequences were isolated at random and then used to found new populations. This process was repeated 4 times until 16 different ancestral DNA sequences had been generated. A rough diagram illustrating the process is shown in Figure 1 on page 2 of the paper.
Now this might not sound like much… but the number of possible phylogenetic trees that can be generated for a group of N different related genetic sequences increases in a steeply exponential manner as N increases. That number is described by the equation: (2N-3)!/((2^(N-2)) (N-2)!).
For 2 organisms this gives us only 1 possible tree (which should be obvious).
For 3 organisms it gives us 3 possible trees.
For 5 it gives us 105.
For 10 it gives us over 34 million.
For 16 organisms that gives us a total of (29!)/((2^14)(14!)) = 29!/1.428x10^15 = 6.19028x10^15 possible phylogenetic tree diagrams that can be generated. Picking the correct one isn’t something you can do by luck... unless of course you can beat better than 6 quintillion to 1 odds. And if that's the case, why aren't you in Vegas right now?
When actually constructing a consensus phylogenetic tree such as the one shown at (Life on Earth) not only are a great many genetic traits taken into account, but a rigorous mathematical analysis of the actual DNA sequences of the organisms in question (where such DNA is available) is done to create cladograms (the branching diagrams showing patterns of descent) with the highest possible percentage confidence. These techniques have been tested in situations where the correct evolutionary relationships are already independently known for an absolute certainty to verify that they do in fact not simply produce an evolutionary relationship but the correct evolutionary relationship to within a very low margin of error..
One example:
http://mbe.oxfordjournals.org/cgi/reprint/19/2/170.pdf
In the paper above the researchers started with an original sample of DNA from Trypanosoma cruzi. They bred it over successive generations and allowed it to continually mutate, and every 70 generations 2 of the resulting DNA sequences were isolated at random and then used to found new populations. This process was repeated 4 times until 16 different ancestral DNA sequences had been generated. A rough diagram illustrating the process is shown in Figure 1 on page 2 of the paper.
Now this might not sound like much… but the number of possible phylogenetic trees that can be generated for a group of N different related genetic sequences increases in a steeply exponential manner as N increases. That number is described by the equation: (2N-3)!/((2^(N-2)) (N-2)!).
For 2 organisms this gives us only 1 possible tree (which should be obvious).
For 3 organisms it gives us 3 possible trees.
For 5 it gives us 105.
For 10 it gives us over 34 million.
For 16 organisms that gives us a total of (29!)/((2^14)(14!)) = 29!/1.428x10^15 = 6.19028x10^15 possible phylogenetic tree diagrams that can be generated. Picking the correct one isn’t something you can do by luck... unless of course you can beat better than 6 quintillion to 1 odds. And if that's the case, why aren't you in Vegas right now?
5.29.2010
Proving Evolution : Post 5 - Non Coding Genetic Sequences
Alright… so we’ve covered radiometric dating and why it’s considered reliable, the geologic column and the fossil record conforming to overall evolutionary expectations, the existence of transitional sequences within the fossil record showing evidence of past evolutionary events, and the distribution of genetic characteristics among modern life that conforms to the pattern produced by a biological evolutionary process in which traits are inherited from common ancestry. We’ve covered that the fossil record also overlays that distribution to a high degree of accuracy with characteristics in inner nested groups in the hierarchy having their first representations later in the fossil record.
Next piece of evidence. Vestigial and other non-coding genetic characteristics.
Vestigial genetic sequences.
Looking at the nested hierarchy shown in the third image of post 4 in this series we can see humans and chimps (along with the rest of the primates) are grouped inside a larger group of animals. Their grouping also indicates a recent evolutionary divergence from that group. This is corroborated by the fossil record. Now… the members of this larger group of animals are capable of synthesizing ascorbic acid, also known as vitamin C. Humans and primates are not. As our little evolutionary branch of the tree only recently diverged from the rest of the group, and since large scale gene deletions are extremely rare (usually a gene is disable because of a disabling mutation… it is not completely removed from the DNA) if evolutionary theory is correct we should expect to still be able to find clear evidence of the genetic sequence responsible for the synthesis of ascorbic acid in humans and primates (even though we are not capable of such synthesis) and subsequently compare it to the functional sequence in other animals and determine what alteration made to it caused it to become non-functional. This is a prediction unique to evolution, relying entirely on the premise that we inherited our genetic material from an ancestral source we share in common with those other animals in the larger group.
This prediction was confirmed in the early 1990s with the identification of the L-gulano-gamma-lactone oxidase genetic code in humans and primates. Subsequent analysis showed it had experienced a frame shift mutation that had caused it to become non-coding.
Let me summarize this again to ensure it is fully understood.
1. Humans and primates do not produce their own ascorbic acid. From simple direct observation there is NO reason to think they would have the genetic code required to do so.
2. The nested hierarchical structure humans and primates fit into within an evolutionary framework however indicates that they diverged from a wider group at a time when ascorbic acid synthesis was already present in the genome of the group, and thus that genetic information should have been inherited.
3. Since we do not produce ascorbic acid, and since it would be unusual to have an entire gene simply deleted in entirety from the genome, evolutionary theory and evolutionary theory alone predicts we should find vestigial genetic code for the production of ascorbic acid which was inherited from an earlier common ancestor in the human and primate genomes… and which has since been deactivated by mutation.
4. They looked for it. They found it. Deactivated by a frame shift mutation that wiped out the end of the sequence on that gene. Prediction confirmed.
Once again… I can’t stop someone from looking at this clear example of evidence of common evolutionary descent and declaring “it just looks that way because it was designed that way” but at this point, whether it’s impossible to disprove that statement or not, it would be beginning to get silly… proposing that the same non functional section of genetic code would be designed into humans and primates… and in such a way that it looked just like a functional piece of code in other animals that had undergone a mutation. If you want to design an organism that doesn’t synthesize its own ascorbic acid you sure as heck don’t need to give it most of the genetic code to do so only to make it not do so!
And this is hardly the only example of a vestigial genetic sequence that fit this pattern…. Olfactory receptor genes, RT6 protein genes, etc… the genetic code of all kind of organisms is packed with pseudogenes that used to code for something in an ancestor… still codes for that same function in related organisms, but has been disabled in one particular group by a crippling mutation while the bulk of the genetic code remains present.
Continuing on that line, there is also the matter of endogenous retroviral insertions.
Next piece of evidence. Vestigial and other non-coding genetic characteristics.
Vestigial genetic sequences.
Looking at the nested hierarchy shown in the third image of post 4 in this series we can see humans and chimps (along with the rest of the primates) are grouped inside a larger group of animals. Their grouping also indicates a recent evolutionary divergence from that group. This is corroborated by the fossil record. Now… the members of this larger group of animals are capable of synthesizing ascorbic acid, also known as vitamin C. Humans and primates are not. As our little evolutionary branch of the tree only recently diverged from the rest of the group, and since large scale gene deletions are extremely rare (usually a gene is disable because of a disabling mutation… it is not completely removed from the DNA) if evolutionary theory is correct we should expect to still be able to find clear evidence of the genetic sequence responsible for the synthesis of ascorbic acid in humans and primates (even though we are not capable of such synthesis) and subsequently compare it to the functional sequence in other animals and determine what alteration made to it caused it to become non-functional. This is a prediction unique to evolution, relying entirely on the premise that we inherited our genetic material from an ancestral source we share in common with those other animals in the larger group.
This prediction was confirmed in the early 1990s with the identification of the L-gulano-gamma-lactone oxidase genetic code in humans and primates. Subsequent analysis showed it had experienced a frame shift mutation that had caused it to become non-coding.
Let me summarize this again to ensure it is fully understood.
1. Humans and primates do not produce their own ascorbic acid. From simple direct observation there is NO reason to think they would have the genetic code required to do so.
2. The nested hierarchical structure humans and primates fit into within an evolutionary framework however indicates that they diverged from a wider group at a time when ascorbic acid synthesis was already present in the genome of the group, and thus that genetic information should have been inherited.
3. Since we do not produce ascorbic acid, and since it would be unusual to have an entire gene simply deleted in entirety from the genome, evolutionary theory and evolutionary theory alone predicts we should find vestigial genetic code for the production of ascorbic acid which was inherited from an earlier common ancestor in the human and primate genomes… and which has since been deactivated by mutation.
4. They looked for it. They found it. Deactivated by a frame shift mutation that wiped out the end of the sequence on that gene. Prediction confirmed.
Once again… I can’t stop someone from looking at this clear example of evidence of common evolutionary descent and declaring “it just looks that way because it was designed that way” but at this point, whether it’s impossible to disprove that statement or not, it would be beginning to get silly… proposing that the same non functional section of genetic code would be designed into humans and primates… and in such a way that it looked just like a functional piece of code in other animals that had undergone a mutation. If you want to design an organism that doesn’t synthesize its own ascorbic acid you sure as heck don’t need to give it most of the genetic code to do so only to make it not do so!
And this is hardly the only example of a vestigial genetic sequence that fit this pattern…. Olfactory receptor genes, RT6 protein genes, etc… the genetic code of all kind of organisms is packed with pseudogenes that used to code for something in an ancestor… still codes for that same function in related organisms, but has been disabled in one particular group by a crippling mutation while the bulk of the genetic code remains present.
Continuing on that line, there is also the matter of endogenous retroviral insertions.
Proving Evolution: Post 4 - The Nested Hierarchy
This is one of the most fundamental concepts which need to be understood if you want to properly grasp the evidence for evolution. It describes the structure of the pattern of biological diversity produced by an evolutionary process. Evolution is the only process ever proposed which predicts and explains such a pattern.
The nested hierarchy is a consequence of the way in which genetically heritable traits are transmitted from generation to generation. For an illustration, see the following:

“NGT” represents a point at which a new genetic trait is introduced to a population which through natural selection comes to be spread throughout that population to the point where it reaches fixation in the genome. From that point on it will be heritable by all the future generations of that population. Such an event can occur at any time, in any group, but due to the nature of biological reproduction and genetic heritability it can be propagated only “downstream” of the point at which it is introduced. So, the earlier in the process a new trait is acquired, the wider a cross-section of the final population it will be present in. Any traits acquired after that point will be found grouped into smaller and smaller cross-sections of the population and always completely contained within the groupings of earlier acquired traits.
For example, let's say that trait "D" up there was the introduction of, say, a specific alteration to the eye of one species that significantly improved it's visual acuity. That species up top on the other hand, developing along that path that leads to them having traits "A" and "F"... their eyes suck. Well, they're out of luck. Because that new trait for improved vision is only able to be passed on through inheritance so they're not getting it. It can only propagate downstream in the process.
To contrast, if we were examining the products of a common design process we not only could, but would expect to see such outcomes all the time. If while working it’s way along those branching design paths a designer came up at some point with that “D” trait and noticed… “hey, this works better than anything I’m using over on that “AF” development line” then of course any designer would utilize that knowledge in his other designs. For example: The human eye vs. the octopus eye. The eyes on an octopus are far superior in design to our eyes. Their optic nerves attach in a manner which does not produce a blind spot in their vision, that same attachment anchors their retinas, and because all the nerve connections come in through the rear of the eye they do not degrade visual acuity. In our eyes the optics nerve pokes through the back of the eye causing a blind spot which our brains must constantly imperfectly correct for, because our retinas are not anchored by the nerve attachments a sharp blow to the head can detach them, and because the nerve attachments are in the front they get in the way of incoming light screwing up our visual acuity. Any designer who knew how to build an octopus eye would know there was a better way to design an eye than that. The evidence indicates however that the octopus evolutionary path simply experienced optical development which was superior to anything that occurred in human ancestry after the two branched off from each other… and after that branching occurred there was no way in which to share the advances experienced on one line with the other. Not within an evolutionary framework.
Now, what do we see when we look at the pattern of biological diversity present today? Here’s an example using 30 major examples (animated gif, modified from the universal phylogenetic tree diagram in Doug Theobald’s “29 evidences for macroevolution”. Takes a little while to cycle through.)
The nested hierarchy is a consequence of the way in which genetically heritable traits are transmitted from generation to generation. For an illustration, see the following:
“NGT” represents a point at which a new genetic trait is introduced to a population which through natural selection comes to be spread throughout that population to the point where it reaches fixation in the genome. From that point on it will be heritable by all the future generations of that population. Such an event can occur at any time, in any group, but due to the nature of biological reproduction and genetic heritability it can be propagated only “downstream” of the point at which it is introduced. So, the earlier in the process a new trait is acquired, the wider a cross-section of the final population it will be present in. Any traits acquired after that point will be found grouped into smaller and smaller cross-sections of the population and always completely contained within the groupings of earlier acquired traits.
For example, let's say that trait "D" up there was the introduction of, say, a specific alteration to the eye of one species that significantly improved it's visual acuity. That species up top on the other hand, developing along that path that leads to them having traits "A" and "F"... their eyes suck. Well, they're out of luck. Because that new trait for improved vision is only able to be passed on through inheritance so they're not getting it. It can only propagate downstream in the process.
To contrast, if we were examining the products of a common design process we not only could, but would expect to see such outcomes all the time. If while working it’s way along those branching design paths a designer came up at some point with that “D” trait and noticed… “hey, this works better than anything I’m using over on that “AF” development line” then of course any designer would utilize that knowledge in his other designs. For example: The human eye vs. the octopus eye. The eyes on an octopus are far superior in design to our eyes. Their optic nerves attach in a manner which does not produce a blind spot in their vision, that same attachment anchors their retinas, and because all the nerve connections come in through the rear of the eye they do not degrade visual acuity. In our eyes the optics nerve pokes through the back of the eye causing a blind spot which our brains must constantly imperfectly correct for, because our retinas are not anchored by the nerve attachments a sharp blow to the head can detach them, and because the nerve attachments are in the front they get in the way of incoming light screwing up our visual acuity. Any designer who knew how to build an octopus eye would know there was a better way to design an eye than that. The evidence indicates however that the octopus evolutionary path simply experienced optical development which was superior to anything that occurred in human ancestry after the two branched off from each other… and after that branching occurred there was no way in which to share the advances experienced on one line with the other. Not within an evolutionary framework.
Now, what do we see when we look at the pattern of biological diversity present today? Here’s an example using 30 major examples (animated gif, modified from the universal phylogenetic tree diagram in Doug Theobald’s “29 evidences for macroevolution”. Takes a little while to cycle through.)
5.28.2010
Proving Evolution: Post 3 - Transitional Fossils
Having covered in posts 1 and 2 why radiometric dating is considered reliable, how the geologic column appears, and how the cross correlation of radiometric dates, fossil composition, and layer depth in the column all converge on expectations it’s time to take a closer look at some of the transitional sequences in the fossil record. The Transitional Vertbrate Fossils FAQ at talk.origins does it very well so I’ll leave the listing to them:
Instead, here I’ll focus on just a few example transitions with some detailed discussion.
Reptiles to Mammals
The list provided at the link above for this particular transition is extensive, covering a sequence of 30 fossil species… the early quite reptilian, then reptilian but with some somewhat mammalian features… then reptilian with some more than somewhat mammalian features… then a solid mix of reptilian and mammalian features… then decidedly mammalian with reptilian features… then mammalian with some somewhat reptilian features, and finally mammalian with few if any reptilian features.
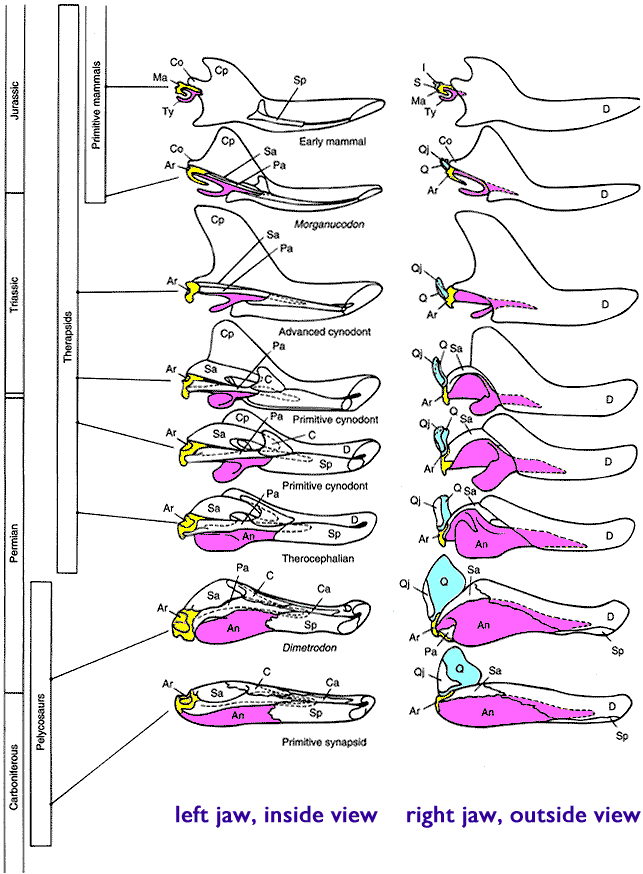
One particularly well illustrated example they proivide of what was occurring during this process is the development of the mammalian ear from reptile jaw structures… illustration here:
Starting at the far left side of the image we have the timescale of which periods each of the fossils are found from. As we move forward from the Carboniferous to the Jurassic we see the clear gradual change in the shape of the skeletal structure in each consecutive example. The left hand column of images if the view of the jaw from the inside. The right hand column is the view of the same jaw from the outside. The bone highlighted in yellow is the articular reptilian jaw bone, which eventually becomes the mammalian malleus (the “hammer” in the ear). The bone hignlighted in pink is the reptilian angular jaw bone, which eventually becomes the tympanic annulus in mammals. The bone highlighted in light blue is the reptilian quadrate jaw bone, which eventually becomes the mammalian incus (the “anvil” in the ear).
This particular sequence is also an excellent illustration of the gaping flaw in claims of “Irreducible Complexity”. Such arguments simply don’t understand how evolution progresses. Someone who held to the IC line of argument would look at something like the ear and say “Well, what good is an ear without the hammer? Huh? What good is half an ear? All those interconnecting bones would have to evolve all at the same time! That’s just silly... so the ear is Irreducibly Complex”. (And yes, people really do make that argument)
On the surface of it, if you don’t really understand how evolution operates, that article has a certain compelling appeal to common sense. People who haven’t been exposed to the full weight of the evidence for evolution and how it operates think that is a perfectly reasonable statement. It is however dead wrong, as we can clearly see. They look at a modern human ear, which is the product of millions of years of refinement to optimize it for operating with the structures available to it… and then just because after all that fine tuning if you suddenly come along and yank a gear out of the mechanism it stops working it couldn’t have developed gradually? Nonsense. Nobody with any knowledge of evolution would ever say that at some point in the past there was some animal with an ear that was completely missing a malleus… but otherwise was an ear exactly like a modern human with all the same bones in the same shape for no apparent reason whatsoever… just waiting around for a fluke mutation to pop that bone right in there out of nowhere. That is an absurd representation of evolutionary progression and does not remotely resemble what is encompassed by evolutionary theory. It is nothing but a flimsy strawman.
On to the next example:
Proving Evolution : Post 2 - The Geologic Column
When the geologic column was first being mapped out by geologists they could only establish relative dates of the position of formation of a given layer in the column based on the premise that 'layers buried further down' = 'older than newly formed surface layers'… with care being taken to ensure you weren’t analyzing something like an overthrust where one section of plate has pushed up on top of another one. Then came radiometric dating which allowed them not only to independently test that hypothesis but to assign specific age values to each of those layers… resulting in the modern understanding of the geologic column. For example, in Glenn Morton’s article on the geologic column at Talk.Origins (The Entire Geologic Column in North Dakota) one of the references used is a well dug in North Dakota to a depth of over 15 thousand feet. The following layers were encountered at the respective depths: (Fm = Formation, Lm = Limestone, Grp = Group)
Tertiary Ft. Union Fm ...............................100 feet
Cretaceous Greenhorn Fm .......................4910 feet
Cretaceous Mowry Fm............................ 5370 feet
Cretaceous Inyan Kara Fm.......................5790 feet
Jurassic Rierdon Fm................................6690 feet
Triassic Spearfish Fm..............................7325 feet
Permian Opeche Fm................................7740 feet
Pennsylvanian Amsden Fm.......................7990 feet
Pennsylvanian Tyler Fm...........................8245 feet
Mississippian Otter Fm.............................8440 feet
Mississippian Kibbey Lm...........................8780 feet
Mississippian Charles Fm..........................8945 feet
Mississippian Mission Canyon Fm................9775 feet
Mississippian Lodgepole Fm.....................10255 feet
Devonian Bakken Fm.............................11085 feet
Devonian Birdbear Fm............................11340 feet
Devonian Duperow Fm...........................11422 feet
Devonian Souris River Fm.......................11832 feet
Devonian Dawson Bay Fm.......................12089 feet
Devonian Prairie Fm...............................12180 feet
Devonian Winnipegosis Grp.....................12310 feet
Silurian Interlake Fm..............................12539 feet
Ordovician Stonewall Fm........................13250 feet
Ordovician Red River Dolomite.................13630 feet
Ordovician Winnipeg Grp........................14210 feet
Ordovician Black Island Fm.....................14355 feet
Cambrian Deadwood Fm.........................14445 feet
Precambrian.........................................14945 feet
The article also includes 25 other sites where the entire column has been observed.
The span of ages since associated with each of those eras since the advent of radiometric dating are:
Tertiary –------------------ 1.8 million -> 65 million years old
Cretaceous --------------– 65 million -> 145 million years old
Jurassic ----------------– 145 million -> 205 million years old
Triassic ----------------–205 million -> 250 million years old
Permian –---------------- 250 million -> 290 million years old
Pennsylvanian –---------- 290 million -> 325 million years old
Mississippian –------------ 325 million to 355 million years old
Devonian –--------------- 355 million -> 420 million years old
Silurian –----------------- 420 million -> 445 million years old
Ordovician –-------------- 445 million -> 490 million years old
Cambrian Deadwood Fm –- 490 million -> 545 million years old
Precambrian –------------------------ 545+ million years old
So, the further down we go, the older the dates we see. Exactly as predicted. But that isn’t the only indicator to consider, there is also the fossil composition of the geologic column. I’ll do a quick overview of which fossils are found in which layers for now... starting with what are dated as the oldest layers and progressing through to the youngest. Note that the precise locations of many of these "earliest known fossil" finds are constantly being adjusted to some degree as more and more fossil finds come in and the body of what is known is added to.... for example, not too many years ago the earliest known multicellular fossils were early Cambrian (540 million years old) but then someone found some in layers about 20 million years older than that and the date of the earliest known multicellular fossils got shifted back a few percent into the late Precambrian. This is to be expected... and will certainly continue to happen in the future.
Precambrian
--In the oldest dated layers of rock in the Precambrian there has never been a fossil found. Of anything. Ever.
--As we move to newer layers in the Precambrian we start finding fossils of single celled organisms at about the 3.5 billion year mark. They appear to be prokaryotes. We find fossils of nothing else.
--In still newer layers we begin finding fossils of what seem to be eukaryotic single celled organisms. (Prokaryotes have cell structures that lack mitochondria and nuclei, eukaryotes incorporate mitochondria and nuclei).
--In the late Precambrian layers leading up to the Cambrian, we begin finding fossils of small, simple, multicellular organisms (for example: the Ediacaran fauna) and also fossils of what appear to be simple chloroplasts.
Tertiary Ft. Union Fm ...............................100 feet
Cretaceous Greenhorn Fm .......................4910 feet
Cretaceous Mowry Fm............................ 5370 feet
Cretaceous Inyan Kara Fm.......................5790 feet
Jurassic Rierdon Fm................................6690 feet
Triassic Spearfish Fm..............................7325 feet
Permian Opeche Fm................................7740 feet
Pennsylvanian Amsden Fm.......................7990 feet
Pennsylvanian Tyler Fm...........................8245 feet
Mississippian Otter Fm.............................8440 feet
Mississippian Kibbey Lm...........................8780 feet
Mississippian Charles Fm..........................8945 feet
Mississippian Mission Canyon Fm................9775 feet
Mississippian Lodgepole Fm.....................10255 feet
Devonian Bakken Fm.............................11085 feet
Devonian Birdbear Fm............................11340 feet
Devonian Duperow Fm...........................11422 feet
Devonian Souris River Fm.......................11832 feet
Devonian Dawson Bay Fm.......................12089 feet
Devonian Prairie Fm...............................12180 feet
Devonian Winnipegosis Grp.....................12310 feet
Silurian Interlake Fm..............................12539 feet
Ordovician Stonewall Fm........................13250 feet
Ordovician Red River Dolomite.................13630 feet
Ordovician Winnipeg Grp........................14210 feet
Ordovician Black Island Fm.....................14355 feet
Cambrian Deadwood Fm.........................14445 feet
Precambrian.........................................14945 feet
The article also includes 25 other sites where the entire column has been observed.
The span of ages since associated with each of those eras since the advent of radiometric dating are:
Tertiary –------------------ 1.8 million -> 65 million years old
Cretaceous --------------– 65 million -> 145 million years old
Jurassic ----------------– 145 million -> 205 million years old
Triassic ----------------–205 million -> 250 million years old
Permian –---------------- 250 million -> 290 million years old
Pennsylvanian –---------- 290 million -> 325 million years old
Mississippian –------------ 325 million to 355 million years old
Devonian –--------------- 355 million -> 420 million years old
Silurian –----------------- 420 million -> 445 million years old
Ordovician –-------------- 445 million -> 490 million years old
Cambrian Deadwood Fm –- 490 million -> 545 million years old
Precambrian –------------------------ 545+ million years old
So, the further down we go, the older the dates we see. Exactly as predicted. But that isn’t the only indicator to consider, there is also the fossil composition of the geologic column. I’ll do a quick overview of which fossils are found in which layers for now... starting with what are dated as the oldest layers and progressing through to the youngest. Note that the precise locations of many of these "earliest known fossil" finds are constantly being adjusted to some degree as more and more fossil finds come in and the body of what is known is added to.... for example, not too many years ago the earliest known multicellular fossils were early Cambrian (540 million years old) but then someone found some in layers about 20 million years older than that and the date of the earliest known multicellular fossils got shifted back a few percent into the late Precambrian. This is to be expected... and will certainly continue to happen in the future.
Precambrian
--In the oldest dated layers of rock in the Precambrian there has never been a fossil found. Of anything. Ever.
--As we move to newer layers in the Precambrian we start finding fossils of single celled organisms at about the 3.5 billion year mark. They appear to be prokaryotes. We find fossils of nothing else.
--In still newer layers we begin finding fossils of what seem to be eukaryotic single celled organisms. (Prokaryotes have cell structures that lack mitochondria and nuclei, eukaryotes incorporate mitochondria and nuclei).
--In the late Precambrian layers leading up to the Cambrian, we begin finding fossils of small, simple, multicellular organisms (for example: the Ediacaran fauna) and also fossils of what appear to be simple chloroplasts.
5.27.2010
Proving Evolution: Post 1 - Dating Methods
First, a note on the title of the series of posts. Science does NOT "prove" things. Anything. Ever. (No, really... not anything). "Proof" is for math and alcohol. What we are really talking about here is "evidentially supported to the greatest extent manageable". But if you say that then people who don't understand science just declare "Aha! But you didn't PROVE it!" ...as if that was relevant. So, we end up with titles like this. Any science purists out ther who are outraged at the inexact terminology, my abject apologies.
Now, getting down to things. This is a series of posts I wrote up a long, long time ago while banging my head against the brick wall that is "people on the internet who believe in Creationism". I think it does a pretty decent job of covering a LOT of information while maintaining a balance between being not so technical the average non-scientific type can't understand it if they take a real shot at it... but technical enough that it remains accurate and not distorted by horrendous over-simplification. I'll be throwing them up here over he next week or so.
====================================
Dating Methods
Carbon (C14) Dating:
C14 dating is used to date the remains of organic, air breathing organisms up to approximately 50,000 years old. While living these organisms breathe the atmosphere, which contains trace amounts of the radioactive isotope Carbon 14 that is constantly being produced in the upper atmosphere through neutron bombardment. So long as they are alive the C14 content of their bodies will remain in equilibrium with the C14 content of the atmosphere. When they die respiration ceases, along with the intake of any new quantities of C14. Over time the C14 decays with a half-life of 5568 years into N14. By measuring how much C14 remains un-decayed the time elapsed since the death of the organism can be determined.
A common misperception of C14 dating is that it relies on the assumption that atmospheric C14 levels remained constant in the past so that we can know how much C14 an organism started off with. While this was an assumption made when the technique was first developed about half a century ago it has not been the case for several decades. Historical atmospheric C14 concentrations have been charted and calibrated using both dendochronology and lake varves which incorporate organic sediment in their annual deposition layers. One particularly good example of this is Lake Suigetsu in Japan where cores have been drilled to a depth of 45,000 annual layers. Because of the layering process we have an independent count of exactly how old every layer is… and because the layers incorporate organic material (the remains of a surface algae which dies off every year and sinks to the bottom of the lake) each layer can be C14 dated as well, and using these two data points the atmospheric C14 content can be charted all the way back for the entire time span encompassed by the varve core. This data (cross-checked against multiple other sites and methods) then allows us to apply C14 dating to other sites already knowing how fluctuations in atmospheric C14 concentrations in the past will effect the results… and allowing us to calibrate out error that would otherwise be introduced due to those past fluctuations.
Now, getting down to things. This is a series of posts I wrote up a long, long time ago while banging my head against the brick wall that is "people on the internet who believe in Creationism". I think it does a pretty decent job of covering a LOT of information while maintaining a balance between being not so technical the average non-scientific type can't understand it if they take a real shot at it... but technical enough that it remains accurate and not distorted by horrendous over-simplification. I'll be throwing them up here over he next week or so.
====================================
Dating Methods
Carbon (C14) Dating:
C14 dating is used to date the remains of organic, air breathing organisms up to approximately 50,000 years old. While living these organisms breathe the atmosphere, which contains trace amounts of the radioactive isotope Carbon 14 that is constantly being produced in the upper atmosphere through neutron bombardment. So long as they are alive the C14 content of their bodies will remain in equilibrium with the C14 content of the atmosphere. When they die respiration ceases, along with the intake of any new quantities of C14. Over time the C14 decays with a half-life of 5568 years into N14. By measuring how much C14 remains un-decayed the time elapsed since the death of the organism can be determined.
A common misperception of C14 dating is that it relies on the assumption that atmospheric C14 levels remained constant in the past so that we can know how much C14 an organism started off with. While this was an assumption made when the technique was first developed about half a century ago it has not been the case for several decades. Historical atmospheric C14 concentrations have been charted and calibrated using both dendochronology and lake varves which incorporate organic sediment in their annual deposition layers. One particularly good example of this is Lake Suigetsu in Japan where cores have been drilled to a depth of 45,000 annual layers. Because of the layering process we have an independent count of exactly how old every layer is… and because the layers incorporate organic material (the remains of a surface algae which dies off every year and sinks to the bottom of the lake) each layer can be C14 dated as well, and using these two data points the atmospheric C14 content can be charted all the way back for the entire time span encompassed by the varve core. This data (cross-checked against multiple other sites and methods) then allows us to apply C14 dating to other sites already knowing how fluctuations in atmospheric C14 concentrations in the past will effect the results… and allowing us to calibrate out error that would otherwise be introduced due to those past fluctuations.
5.26.2010
"Faith"
Well, I got started with the content creation then kind of slacked off for a while. To try and kick start things again I've hunted down some previous writings of mine from various discussion forums and will be putting them up here.
This one is on a subject that just recently came up again in another conversation I was having... which caused me to remember this post. I wrote this about two and a half years ago... my views on the subject are unchanged since then.
=========================
This is a topic I've wanted to address in greater detail for some time now. Let's start with a few examples of what "faith" is, as there are many different kinds.
1. "Faith" as a description of confidence in extremely reliable data. For example, I have faith the sun will rise tomorrow morning. The reason I have faith in this is because I have massive amounts of data available to me that inform me that this will occur barring an incredibly unlikely occurrence... like the sun exploding overnight or the space fairies halting the Earth's rotation. The sun has risen all 11,000+ days of my life to date, right on schedule, and I have no reason to suspect that pattern will be disrupted in the next 14 hours or so.
2. "Faith" as a description of well earned trust. For example, you can have faith that a good friend or close acquaintance will deal honestly and fairly with you. This is based on your experience of and familiarity with this person and their personality and behavior. Your judgment of their character. Really, you are expressing confidence in your own ability to evaluate the trustworthiness of another person when you encounter and interact with them.
3. "Faith" as an expresssion of loyalty and commitment. Usually to an individual or ideal which you have good reason to hold as worthy of support, as in 'keeping the faith'. You have evaluated and judged this person or principle and have come to the conclusion that it is worthy of your loyalty and efforts to advance it, and faithfully stand by it.
4. "Faith" as a description of an insistence on believing in something without regard for or even in direct opposition to any related information or evidence. For example, to cite some extreme cases, you can have faith that there is a spaceship carrying Jesus riding along and hiding behind the Hale-Bopp comet and if you commit suicide while it passes your soul will float up to zoom around with Jesus in outer space. Or, you can have faith that food and water are unnecessary for your survival, and humans can survive by being photosynthetic or something. People clinging to this type of faith can usually be identified by statements such as "It doesn't matter what you say you can't change my mind, I hold my position through faith and my faith is unshakable!"
This one is on a subject that just recently came up again in another conversation I was having... which caused me to remember this post. I wrote this about two and a half years ago... my views on the subject are unchanged since then.
=========================
This is a topic I've wanted to address in greater detail for some time now. Let's start with a few examples of what "faith" is, as there are many different kinds.
1. "Faith" as a description of confidence in extremely reliable data. For example, I have faith the sun will rise tomorrow morning. The reason I have faith in this is because I have massive amounts of data available to me that inform me that this will occur barring an incredibly unlikely occurrence... like the sun exploding overnight or the space fairies halting the Earth's rotation. The sun has risen all 11,000+ days of my life to date, right on schedule, and I have no reason to suspect that pattern will be disrupted in the next 14 hours or so.
2. "Faith" as a description of well earned trust. For example, you can have faith that a good friend or close acquaintance will deal honestly and fairly with you. This is based on your experience of and familiarity with this person and their personality and behavior. Your judgment of their character. Really, you are expressing confidence in your own ability to evaluate the trustworthiness of another person when you encounter and interact with them.
3. "Faith" as an expresssion of loyalty and commitment. Usually to an individual or ideal which you have good reason to hold as worthy of support, as in 'keeping the faith'. You have evaluated and judged this person or principle and have come to the conclusion that it is worthy of your loyalty and efforts to advance it, and faithfully stand by it.
4. "Faith" as a description of an insistence on believing in something without regard for or even in direct opposition to any related information or evidence. For example, to cite some extreme cases, you can have faith that there is a spaceship carrying Jesus riding along and hiding behind the Hale-Bopp comet and if you commit suicide while it passes your soul will float up to zoom around with Jesus in outer space. Or, you can have faith that food and water are unnecessary for your survival, and humans can survive by being photosynthetic or something. People clinging to this type of faith can usually be identified by statements such as "It doesn't matter what you say you can't change my mind, I hold my position through faith and my faith is unshakable!"
Subscribe to:
Posts (Atom)

{kind=link}